
Page 13 - 《精细化工)》2023年第10期
P. 13
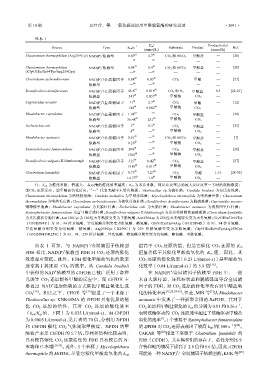
第 10 期 史红玲,等: 二氧化碳还原用甲酸脱氢酶的研究进展 ·2091·
续表 1
K m/ Productivity/
–1
Source Type k cat/s Substrate Product Ref.
(mmol/L) (mmol/h)
①
+
Chaetomium thermophilum (Arg259Cys) NAD(P) 依赖型 0.09 ① 4.7 CO 2 和 HCO 3 – 甲酸盐 — [20]
— — — — —
②
②
①
+
Chaetomium thermophilum NAD(P) 依赖型 0.04 ① 8.4 CO 2 和 HCO 3 – 甲酸盐 — [20]
(Gly93His/Ile94Tyr/Arg259Cys) — — — — —
②
②
Clostridium carboxidivorans NAD(P) /金属辅因子 0.08 ① 0.05 CO 2 甲酸 — [21]
①
+
②
②
依赖型 — — — — —
①
+
Desulfovibrio desulfuricans NAD(P) /金属辅因子 46.6 ① 0.016 CO 2 和 H 2 甲酸盐 0.4 [22-23]
②
②
依赖型 543 0.057 甲酸钠 CO 2 —
①
①
+
Cupriavidus necator NAD(P) /金属辅因子 11 2.7 CO 2 甲酸 — [12]
②
②
依赖型 140 0.082 甲酸钠 CO 2 —
Rhodobacter capsulatus NAD(P) /金属辅因子 1.48 ① — CO 2 甲酸盐 — [24]
+
①
②
依赖型 36.48 ② 281 甲酸钠 CO 2 —
+
Escherichia coli NAD(P) /金属辅因子 1 8.3 CO 2 甲酸盐 — [25]
①
①
②
②
依赖型 4 — 甲酸钠 CO 2 —
①
+
Rhodobacter aestuarii NAD(P) /金属辅因子 0.81 ① — CO 2 和 HCO 3 – 甲酸盐 — [1]
②
依赖型 0.26 ② — 甲酸钠 CO 2 —
+
①
①
Syntrophobacter fumaroxidans NAD(P) /金属辅因子 500 — CO 2 甲酸盐 — [26]
②
依赖型 1500 ② — 甲酸钠 CO 2 —
①
+
①
Desulfovibrio vulgaris Hildenborough NAD(P) /金属辅因子 315 0.42 CO 2 甲酸盐 — [27]
②
依赖型 1310 ② 0.017 甲酸钠 CO 2 —
①
Clostridium ljundahlii NAD(P) /金属辅因子 0.73 ① 7.27 CO 2 甲醇 1.31 [28-30]
+
②
依赖型 14.77 ② 1.4 甲酸钠 CO 2 —
注:k cat 为催化常数,值越大,表示酶的催化效率越高;K m 为米氏常数,即反应速率达到最大反应速率一半时的底物浓度;
①CO 2 还原反应;②甲酸盐氧化反应;“—”代表文献中未给出数据。Thiobacillus sp.为硫杆菌;Candida boidinii 为布氏念珠菌;
Chaetomium thermophilum 为嗜热链球菌;Candida methylica 为甲基念珠菌;Myceliophthora thermophile 为嗜热菌丝体;Chaetomium
thermophilum 为嗜热毛壳菌;Clostridium carboxidivorans 为梭状芽孢杆菌;Desulfovibrio desulfuricans 为脱硫弧菌;Cupriavidus necator
铜绿假单胞菌;Rhodobacter capsulatus 为荚膜红杆菌;Escherichia coli 为大肠杆菌;Rhodobacter aestuarii 为埃斯图里红杆菌;
Syntrophobacter fumaroxidans 为富马酸合成杆菌;Desulfovibrio vulgaris Hildenborough 为希尔登伯勒普通硫弧菌;Clostridium ljundahlii
为艾氏梭状芽胞杆菌。Asn120Cys 为 120 位天冬酰胺突变为半胱氨酸,Asn256Asp 为 256 位天冬酰胺突变为天冬氨酸;Gly93His/Ile94Tyr
(G93H/I94Y)为 93、94 位甘氨酸、异亮氨酸分别突变为组氨酸、酪氨酸;Gly93His/Ile94Arg(G93H/I94R)为 93、94 位甘氨酸、
异亮氨酸分别突变为组氨酸、精氨酸;Arg259Cys(R259C)为 259 位精氨酸突变为半胱氨酸;Gly93His/Ile94Tyr/Arg259Cys
(G93H/I94Y/R259C)为 93、94、259 位甘氨酸、异亮氨酸、精氨酸分别突变为组氨酸、酪氨酸、半胱氨酸。
+
由表 1 可知,与 NAD(P) /金属辅因子依赖型 值高于 CO 2 还原的值,但是它催化 CO 2 还原的 K m
+
FDH 相比,NAD(P) 依赖型 FDH 对 CO 2 还原的催化 值显著低于其催化甲酸盐氧化的 K m 值,因此,其
效率相对较低。此外,FDH 催化甲酸盐氧化的效率 CO 2 还原的催化效率〔0.23 L/(mmols)〕是甲酸盐氧
通常高于其还原 CO 2 的效率。从 Candida boidinii 化效率〔0.04 L/(mmols)〕的 5.8 倍 [18] 。
+
+
中获得的 NAD 依赖型的 CbFDH 已被广泛用于各种 在 NAD(P) /金属辅因子依赖型 FDH 中,一些
光诱导 CO 2 还原制备甲酸的反应中,而 CbFDH 主 来自大肠杆菌、坏死杯状菌和脱硫弧菌等含金属辅
+
要通过 NAD 还原偶联的方式催化甲酸盐氧化生成 因子的 FDH,对 CO 2 还原的转化率没有对甲酸盐氧
[16] 。相比之下,CHOE 等 [16] 报道了一个来源于 化的转化率高 [12,22,24-25] 。但是,MIN 等 从 Rhodobacter
[1]
CO 2
Thiobacillus sp. KNK65MA 的 TsFDH 具有优异的催 aestuarii 中发现了一种新型含钼的 RaFDH,其对于
–1
化 CO 2 还原的活性,其对 CO 2 还原的催化效率 CO 2 还原和甲酸盐氧化的 k cat 值分别为 0.81 和 0.26 s ,
(k cat/K m 值,下同)为 0.035 L/(mmols),而 CbFDH 表明该酶驱动的 CO 2 还原速率超过了该酶驱动甲酸盐
[1]
为 0.0005 L/(mmols),是后者的 70 倍。分别用 TsFDH 氧化的速率 。1 个来源于 Syntrophobacter fumaroxidans
–1 [26]
和 CbFDH 催化 CO 2 气体制取甲酸盐,TsFDH 的甲 的 SfFDH 对 CO 2 还原表现出了较高 k cat 值(500 s ) 。
酸盐产率是 CbFDH 的 5.7 倍。序列和结构比较表明, ÇAKAR 等 [28] 报道了来源于 Clostridium ljundahlii 的
具有较高催化 CO 2 还原活性的 FDH 具有较长的 N- FDH(ClFDH),其在钨和钼的存在下,活性分别比不
末端和 C-末端 [16,31] 。此外,1 个来源于 Myceliophthora 含钨和钼的情况下提高了 2.8 倍和 5.6 倍。因此,ClFDH
+
可能是一种 NAD(P) / 金属辅因子依赖型酶。KUK 等 [29]
thermophile 的 MtFDH,尽管它催化甲酸盐氧化的 k cat