
Page 45 - 201904
P. 45
第 4 期 王小艳,等: 淀粉质燃料乙醇发酵胁迫及菌株耐受性改造 ·571·
一胁迫耐受性后,其生产性能大幅降低的主要原因。 胞壁会发生了一系列的重构 [13] ;再如,高糖胁迫下,
在高温、高糖、高乙醇胁迫下微生物的耐受分 质膜和液泡膜上的 ABC 转运蛋白保持胞质中的离
子机制具有很大的重叠性,但针对不同胁迫也具有 子浓度相对稳定 [14] 。由此可见,不同环境胁迫下微
一定的倾向性,如热休克应答系统是耐热的主要调 生物的耐受机制既有重叠又有区别,因此,在单一
控机制,其热胁迫下所诱导的热激蛋白种类及数量 环境胁迫下获得的耐受性很难实现在真实发酵微环
区别于乙醇胁迫 [12] ;又如,在乙醇压力下,酵母细 境多重胁迫下的实际耐受性。
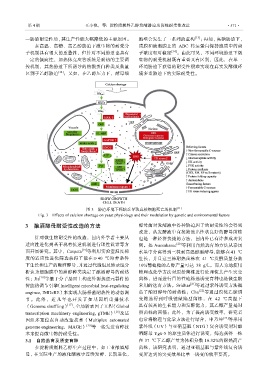
图 3 胁迫环境下钙缺乏导致真核细胞死亡的机制 [11]
Fig. 3 Effects of calcium shortage on yeast physiology and their modulation by genetic and environmental factors
3 酿酒酵母耐受性改造的方法 酵母菌对发酵液中各种胁迫因子的耐受性均会得到
改进,从发酵液中直接筛选出性状良好的酵母菌株
针对微生物耐受性的改造,国内外学者主要从 也是一种经济快捷的方法,国内外已有许多成功案
适应性进化到基于现有抗逆机制进行理性设计等方 例。如 Auesukaree [20] 等利用自然选育的方法从泰国
面开展研究。其中,Caspeta [15] 等利用实验室温度梯 水果中分离得到三株耐高温酿酒酵母,能够在 41 ℃
度的适应性进化筛选获得了能在≥40 ℃培养条件 生长,并且这三株耐热菌株在 41 ℃发酵质量分数
下生长和生产的酿酒酵母,并通过代谢组及转录组分 10%葡萄糖的乙醇产量可达 38 g/L。而人为地使用
析认为细胞膜中的甾醇种类决定了酿酒酵母的耐热 物理或化学方法对原始菌株进行处理使其产生突变
性;Jia [16] 等基于分子层面上构建外源抗逆元器件的 菌株,进而进行目的性地筛选诱变育种也是微生物
智能热调节引擎(Intelligent microbial heat-regulating 常用的选育方法。Sridhar [21] 等通过紫外诱变方法提
enginee, IMHeRE)来实现大肠杆菌耐热性的动态调 高了酿酒酵母的耐热性;Cha [22] 等通过溴化乙腚诱
节。此外,近几年也开发了如基因组重排技术 变筛选得到呼吸链缺陷型菌株,在 42 ℃高温下
[17]
(Genome shuffling) 、全局转录因子工程(Global 具有较高的生长能力和发酵能力,其乙醇产量是对
transcription machinery engineering, gTME) [18] 及基 照菌株的两倍;此外,为了提高诱变效率,研究者
因组多重位点自动改造技术(Multiplex automated 也常将物理与化学方法进行结合,申乃坤 [23] 等采用
genome engineering,MAGE) [19] 等一些先进育种技 紫外线(UV)与亚硝基胍(NTG)复合诱变剂对酿
术来提高微生物的耐受性。 酒酵母 Ygx-5 的原生质体进行诱变,筛选获得一株
3.1 自然选育及诱变育种 在 37 ℃下乙醇产量为体积分数 16.32%的耐热高产
在淀粉质燃料乙醇生产过程中,如工业酿酒酵 菌株,该研究表明,通过亚硝基胍与紫外线复合诱
母,在实际生产的液化醪液中连续发酵、长期进化, 变所达到的突变效果比单一诱变的效率要高。